Математическая модель хищник жертва примеры. Модель ситуации типа «хищник-жертва. Модель ситуации типа «хищник-жертва»
Приспособления, вырабатываемые жертвами для противодействия хищникам , способствуют выработке у хищников механизмов преодоления этих приспособлений. Длительное совместное существование хищников и жертв приводит к формированию системы взаимодействия, при которой обе группы устойчиво сохраняются на изучаемой территории. Нарушение такой системы часто приводит к отрицательным экологическим последствиям.
Негативное влияние нарушения коэволюционных связей наблюдается при интродукции видов. В частности, козы и кролики , интродуцированные в Австралии , не имеют на этом материке эффективных механизмов регуляции численности, что приводит к разрушению природных экосистем .
Математическая модель
Допустим, что на некоторой территории обитают два вида животных : кролики (питающиеся растениями) и лисы (питающиеся кроликами). Пусть число кроликов , число лис . Используя Модель Мальтуса с необходимыми поправками, учитывающими поедание кроликов лисами, приходим к следующей системе, носящей имя модели Вольтерры - Лотки :
\begin{cases} \dot x=(\alpha -c y)x;\\
\dot y=(-\beta+d x) y. \end{cases}
Поведение модели
Групповой образ жизни хищников и их жертв радикально меняет поведение модели, придает ей повышенную устойчивость.
Обоснование: при групповом образе жизни снижается частота случайных встреч хищников с потенциальными жертвами, что подтверждается наблюдениями за динамикой численности львов и антилоп гну в парке Серенгети .
История
Модель совместного существования двух биологических видов (популяций) типа «хищник - жертва» называется также моделью Вольтерры - Лотки.
См. также
Напишите отзыв о статье "Система «хищник - жертва»"
Примечания
Литература
- В. Вольтерра, Математическая теория борьбы за существование. Пер. с франц. О. Н. Бондаренко. Под ред и послесловием Ю. М. Свирежева. М.: Наука, 1976. 287 c. ISBN 5-93972-312-8
- А. Д. Базыкин, Математическая биофизика взаимодействующих популяций. М.: Наука, 1985. 181 с.
- А. Д. Базыкин, Ю. А. Кузнецов, А. И. Хибник, Портреты бифуркаций (Бифуркационные диаграммы- динамических систем на плоскости) /Серия «Новое в жизни, науке, технике. Математика, кибернетика» - М.: Знание, 1989. 48 с.
- П. В. Турчин,
Ссылки
Отрывок, характеризующий Система «хищник - жертва»
– Charmant, charmant, [Прелестно, прелестно,] – сказал князь Василий.– C"est la route de Varsovie peut etre, [Это варшавская дорога, может быть.] – громко и неожиданно сказал князь Ипполит. Все оглянулись на него, не понимая того, что он хотел сказать этим. Князь Ипполит тоже с веселым удивлением оглядывался вокруг себя. Он так же, как и другие, не понимал того, что значили сказанные им слова. Он во время своей дипломатической карьеры не раз замечал, что таким образом сказанные вдруг слова оказывались очень остроумны, и он на всякий случай сказал эти слова, первые пришедшие ему на язык. «Может, выйдет очень хорошо, – думал он, – а ежели не выйдет, они там сумеют это устроить». Действительно, в то время как воцарилось неловкое молчание, вошло то недостаточно патриотическое лицо, которого ждала для обращения Анна Павловна, и она, улыбаясь и погрозив пальцем Ипполиту, пригласила князя Василия к столу, и, поднося ему две свечи и рукопись, попросила его начать. Все замолкло.
– Всемилостивейший государь император! – строго провозгласил князь Василий и оглянул публику, как будто спрашивая, не имеет ли кто сказать что нибудь против этого. Но никто ничего не сказал. – «Первопрестольный град Москва, Новый Иерусалим, приемлет Христа своего, – вдруг ударил он на слове своего, – яко мать во объятия усердных сынов своих, и сквозь возникающую мглу, провидя блистательную славу твоея державы, поет в восторге: «Осанна, благословен грядый!» – Князь Василий плачущим голосом произнес эти последние слова.
Билибин рассматривал внимательно свои ногти, и многие, видимо, робели, как бы спрашивая, в чем же они виноваты? Анна Павловна шепотом повторяла уже вперед, как старушка молитву причастия: «Пусть дерзкий и наглый Голиаф…» – прошептала она.
Князь Василий продолжал:
– «Пусть дерзкий и наглый Голиаф от пределов Франции обносит на краях России смертоносные ужасы; кроткая вера, сия праща российского Давида, сразит внезапно главу кровожаждущей его гордыни. Се образ преподобного Сергия, древнего ревнителя о благе нашего отечества, приносится вашему императорскому величеству. Болезную, что слабеющие мои силы препятствуют мне насладиться любезнейшим вашим лицезрением. Теплые воссылаю к небесам молитвы, да всесильный возвеличит род правых и исполнит во благих желания вашего величества».
– Quelle force! Quel style! [Какая сила! Какой слог!] – послышались похвалы чтецу и сочинителю. Воодушевленные этой речью, гости Анны Павловны долго еще говорили о положении отечества и делали различные предположения об исходе сражения, которое на днях должно было быть дано.
– Vous verrez, [Вы увидите.] – сказала Анна Павловна, – что завтра, в день рождения государя, мы получим известие. У меня есть хорошее предчувствие.
Предчувствие Анны Павловны действительно оправдалось. На другой день, во время молебствия во дворце по случаю дня рождения государя, князь Волконский был вызван из церкви и получил конверт от князя Кутузова. Это было донесение Кутузова, писанное в день сражения из Татариновой. Кутузов писал, что русские не отступили ни на шаг, что французы потеряли гораздо более нашего, что он доносит второпях с поля сражения, не успев еще собрать последних сведений. Стало быть, это была победа. И тотчас же, не выходя из храма, была воздана творцу благодарность за его помощь и за победу.
Предчувствие Анны Павловны оправдалось, и в городе все утро царствовало радостно праздничное настроение духа. Все признавали победу совершенною, и некоторые уже говорили о пленении самого Наполеона, о низложении его и избрании новой главы для Франции.
Вдали от дела и среди условий придворной жизни весьма трудно, чтобы события отражались во всей их полноте и силе. Невольно события общие группируются около одного какого нибудь частного случая. Так теперь главная радость придворных заключалась столько же в том, что мы победили, сколько и в том, что известие об этой победе пришлось именно в день рождения государя. Это было как удавшийся сюрприз. В известии Кутузова сказано было тоже о потерях русских, и в числе их названы Тучков, Багратион, Кутайсов. Тоже и печальная сторона события невольно в здешнем, петербургском мире сгруппировалась около одного события – смерти Кутайсова. Его все знали, государь любил его, он был молод и интересен. В этот день все встречались с словами:
– Как удивительно случилось. В самый молебен. А какая потеря Кутайсов! Ах, как жаль!
– Что я вам говорил про Кутузова? – говорил теперь князь Василий с гордостью пророка. – Я говорил всегда, что он один способен победить Наполеона.
Но на другой день не получалось известия из армии, и общий голос стал тревожен. Придворные страдали за страдания неизвестности, в которой находился государь.
– Каково положение государя! – говорили придворные и уже не превозносили, как третьего дня, а теперь осуждали Кутузова, бывшего причиной беспокойства государя. Князь Василий в этот день уже не хвастался более своим protege Кутузовым, а хранил молчание, когда речь заходила о главнокомандующем. Кроме того, к вечеру этого дня как будто все соединилось для того, чтобы повергнуть в тревогу и беспокойство петербургских жителей: присоединилась еще одна страшная новость. Графиня Елена Безухова скоропостижно умерла от этой страшной болезни, которую так приятно было выговаривать. Официально в больших обществах все говорили, что графиня Безухова умерла от страшного припадка angine pectorale [грудной ангины], но в интимных кружках рассказывали подробности о том, как le medecin intime de la Reine d"Espagne [лейб медик королевы испанской] предписал Элен небольшие дозы какого то лекарства для произведения известного действия; но как Элен, мучимая тем, что старый граф подозревал ее, и тем, что муж, которому она писала (этот несчастный развратный Пьер), не отвечал ей, вдруг приняла огромную дозу выписанного ей лекарства и умерла в мучениях, прежде чем могли подать помощь. Рассказывали, что князь Василий и старый граф взялись было за итальянца; но итальянец показал такие записки от несчастной покойницы, что его тотчас же отпустили.
Часто представители одного вида (популяции) питаются представителями другого вида.
Модель Лотки – Вольтерры – модель взаимного существования двух популяций типа «хищник – жертва».
Впервые модель «хищник – жертва» была получена А. Лоткой в 1925 году, который использовал ее для описания динамики взаимодействующих биологических популяций. В 1926 году независимо от Лотки аналогичные (к тому же более сложные) модели были разработаны итальянским математиком В. Вольтерра, глубокие исследования которого в области экологических проблем заложили фундамент математической теории биологических сообществ или т. н. математической экологии.
В математической форме предложенная система уравнений имеет вид:
где x – количество жертв, y – количество хищников, t – время, α, β, γ, δ – коэффициенты, которые отражают взаимодействия между популяциями.
Постановка задачи
Рассмотрим закрытое пространство, в котором существуют две популяции – травоядные («жертвы») и хищники. Считается, что животных не ввозят и не вывозят и что еды для травоядных животных достаточно. Тогда уравнение изменения числа жертв (только жертв) примет вид:
где $α$ – коэффициент рождаемости жертв,
$x$– размер популяции жертв,
$\frac{dx}{dt}$ – скорость прироста популяции жертв.
Когда хищники не охотятся, они могут вымирать, значит, уравнение для количества хищников (только хищников) примет вид:
Где $γ$ – коэффициент убыли хищников,
$y$ – размер популяции хищников,
$\frac{dy}{dt}$ – скорость прироста популяции хищников.
При встрече хищников и жертв (частота встреч прямо пропорциональна произведению) хищники уничтожают жертв с коэффициентом, сытые хищники могут воспроизводить потомство с коэффициентом. Таким образом, система уравнений модели примет вид:
Решение задачи
Построим математическую модель совместного существования двух биологических популяций типа «хищник – жертва».
Пусть две биологические популяции совместно обитают в изолированной среде. Среда является стационарной и обеспечивает в неограниченном количестве всем необходимым для жизни один из видов – жертвы. Другой вид – хищник – также обитает в стационарных условиях, но питается только жертвами. В роли хищников могут выступать коты, волки, щуки, лисы, а в роли жертв – куры, зайцы, караси, мыши соответственно.
Для определенности рассмотрим в роли хищников – котов, а в роли жертв – кур.
Итак, куры и коты живут в некотором изолированном пространстве – хозяйственном дворе. Среда предоставляет курам питание в неограниченном количестве, а коты питаются только курами. Обозначим через
$х$ – количество кур,
$у$ – количество котов.
Со временем количество кур и котов меняется, но будем считать $х$ и $у$ непрерывными функциями от времени t. Назовем пару чисел $х, у)$ состоянием модели.
Найдем каким образом изменяется состояние модели $(х, у).$
Рассмотрим $\frac{dx}{dt}$ – скорость изменения количества кур.
Если котов нет, то количество кур возрастает и тем быстрее, чем больше кур. Будем считать зависимость линейной:
$\frac{dx}{dt} a_1 x$,
$a_1$ – коэффициент, который зависит только от условий жизни кур, их естественной смертности и рождаемости.
$\frac{dy}{dt}$ – скорость изменения количества котов (если нет кур), зависит от количества котов y.
Если кур нет, то количество котов уменьшается (у них нет пищи) и они вымирают. Будем считать зависимость линейной:
$\frac{dy}{dt} - a_2 y$.
В экосистеме скорость изменения количества каждого вида также будем считать пропорциональным его количеству, но только с коэффициентом, зависящим от количества особей другого вида. Так, для кур этот коэффициент уменьшается с увеличением количества котов, а для котов возрастает с увеличением количества кур. Будем считать зависимость также линейной. Тогда получим систему дифференциальных уравнений:
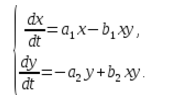
Данная система уравнений называется моделью Вольтерра-Лотки.
a1, a2, b1, b2 – числовые коэффициенты, которые называют параметрами модели.
Как видно, характер изменения состояния модели (x, y) определяется значениями параметров. Изменяя данные параметры и решая систему уравнений модели, можно исследовать закономерности изменения состояния экологической системы.
С помощью программы MATLAB система уравнений Лотки-Вольтерра решается следующим образом:
На рис. 1 представлено решение системы. В зависимости от начальных условий решения разные, чему отвечают разные цвета траекторий.
На рис. 2 представлены те же решения, но с учетом оси времени t (т.е. наблюдается зависимость от времени).
Еще в 20-х гг. А. Лотка (Lotka), а несколько позднее независимо от него В. Вольтерра предложили математические модели, описывающие сопряженные колебания численности популяций хищника и жертвы. Рассмотрим самый простой вариант модели Лотки-Вольтерра. В основе модели лежит ряд допущений:
1) популяция жертв в отсутствие хищника растет экспоненциально,
2) пресс хищников тормозит этот рост,
3) смертность жертв пропорциональна частоте встреч хищника и жертвы (или иначе, пропорциональна произведению плотностей их популяций);
4) рождаемость хищника зависит от интенсивности потребления жертв.
Мгновенная скорость изменения численности популяции жертв может быть выражена уравнением
dN ж /dt = r 1 N ж - p 1 N ж N х,
где r 1 - удельная мгновенная скорость популяционного роста жертвы, p 1 - константа, связывающая смертность жертв с плотностью хищника, a N ж и N х - плотности соответственно жертвы и хищника.
Мгновенная скорость роста популяции хищника в этой модели принимается равной разности рождаемости и постоянной смертности:
dN х /dt = p 2 N ж N х – d 2 N х,
где p 2 - константа, связывающая рождаемость в популяции хищника с плотностью жертв, a d 2 - удельная смертность хищника.
Согласно приведенным уравнениям каждая из взаимодействующих популяций в своем увеличении ограничена только другой популяцией, т.е. рост числа жертв лимитируется прессом хищников, а рост числа хищников - недостаточным количеством жертв. Никакого самоограничения популяций не предполагается. Считается, например, что пищи для жертвы всегда достаточно. Также не предполагается и выхода популяции жертв из-под контроля хищника, хотя на самом деле такое бывает достаточно часто.
Несмотря на всю условность модели Лотки-Вольтерра, она заслуживает внимания уже хотя бы потому, что показывает, как даже такая идеализированная система взаимодействия двух популяций может порождать достаточно сложную динамику их численности. Решение системы этих уравнений позволяет сформулировать условия поддержания постоянной (равновесной) численности каждого из видoв. Популяция жертв сохраняет постоянную численность, если плотность хищника равна r 1 /p 1, а для того чтобы постоянство сохраняла популяция хищника, плотность жертв должна быть равна d 2 /p 2 . Если на графике отложить по оси абсцисс плотность жертв N ж , а по оси ординат - плотность хищника N х , то изоклины, показывающие условие постоянства хищника и жертвы, будут представлять собой две прямые, перпендикулярные друг другу и координатным осям (рис. 6,а). При этом предполагается, что ниже определенной (равной d 2 /p 2) плотности жертв плотность хищника всегда будет уменьшаться, а выше - всегда увеличиваться. Соответственно и плотность жертвы возрастает, если плотность хищника ниже значения, равного r 1 /p 1 , и уменьшается, если, она выше этого значения. Точка пересечения изоклин соответствует условию постоянства численности хищника и жертвы, а другие точки на плоскости этого графика совершают движение по замкнутым траекториям, отражая, таким образом, регулярные колебания численности хищника и жертвы (рис. 6,б). Размах колебаний определяется начальным соотношением плотностей хищника и жертвы. Чем ближе оно к точке пересечения изоклин, тем меньше окружность, описываемая векторами, и соответственно меньше амплитуда колебаний.
Рис. 6. Графическое выражение модели Лотки-Вольтера для системы хищник-жертва.
Одна из первых попыток получения колебаний численности хищника и жертвы в лабораторных экспериментах принадлежала Г.Ф. Гаузе (Gause). Объектами этих экспериментов были инфузория парамеция (Paramecium caudatum ) и хищная инфузория дидиниум (Didinium nasutum ). Пищей для парамеции служила регулярно вносимая в среду взвесь бактерий, а дидиниум питался только парамециями. Данная система оказалась крайне неустойчивой: пресс хищника по мере увеличения его численности приводил к полному истреблению жертв, после чего вымирала и популяция самого хищника. Усложняя опыты, Гаузе устраивал убежище для жертвы, внося в пробирки с инфузориями немного стеклянной ваты. Среди нитей ваты могли свободно перемещаться парамеции, но не могли дидиниумы. В таком варианте опыта дидиниум съедал всех парамеций, плавающих в свободной от ваты части пробирки, и вымирал, а популяция парамеции затем восстанавливалась за счет размножения особей, уцелевших в убежище. Некоторого подобия колебаний численности хищника и жертвы Гаузе удалось добиться только в том случае, когда он время от времени вносил в культуру и жертву и хищника, имитируя таким образом иммиграцию.
Через 40 лет после работы Гаузе его опыты были повторены Л. Лакинбиялом (Luckinbill), использовавшим в качестве жертвы инфузорию Paramecium aurelia , а в качестве хищника того же Didinium nasutum . Лакинбиллу удалось получить несколько циклов колебаний численности этих популяций, но только в том случае, когда плотность парамеций была лимитирована нехваткой пищи (бактерий), а в культуральную жидкость добавляли метилцеллюлозу - вещество, снижающее скорость движения как хищника, так и жертвы и потому уменьшающее частоту их возможных встреч. Оказалось также, что добиться колебаний хищника и жертвы легче, если увеличить объем экспериментального сосуда, хотя условие пищевого лимитирования жертвы и в этом случае обязательно. Если же к системе сосуществующих в колебательном режиме хищника и жертвы добавляли избыточную пищу, то ответом был, быстрый рост численности жертвы, за которым следовало возрастание численности хищника, приводящее в свою очередь к полному истреблению популяции жертвы.
Модели Лотки и Вольтерры послужили толчком для разработки ряда других более реалистичных моделей системы хищник-жертва. В частности, довольно простая графическая модель, анализирующая соотношение разных изоклин жертвы хищника, была предложена М. Розенцвейгом и Р. Мак-Артуров (Rosenzweig, MacArthur). Согласно этим авторам, стационарная (= постоянная) численность жертвы в координатных осях плотности хищника и жертвы может быть представлена в виде выпуклой изоклины (рис. 7,а). Одна точка пересечения изоклины сосью плотности жертвы соответствует минимальной допустимой плотности жертвы (нижеее популяция подвержена очень большому риску вымирания хотя бы из-за малой частоты встреч самцов и самок), а другая - максимальной, определяемой количеством имеющейся пищи или поведенческими особенностями самой жертвы. Подчеркнем, что речь идет пока о минимальной и максимальной плотностях в отсутствие хищника. При появлении хищника и увеличении его численности минимальная допустимая плотность жертвы, очевидно, должна быть выше, а максимальная - ниже. Каждому значению плотности жертвы должна соответствовать некоторая плотность хищника, при которой достигается постоянство популяции жертвы. Геометрическое место таких точек и есть изоклина жертвы в координатах плотности хищника и жертвы. Векторы, показывающие направление изменения плотности жертвы (ориентированные горизонтально), имеют разную направленность по разные стороны от изоклины (рис. 7,а).

Рис. 7. Изоклины стационарных популяций жертвы (а) и хищника (б).
Для хищника в тех же координатах также построена изоклина, отвечающая стационарному состоянию его популяции. Векторы, показывающие направление изменения численности хищника, ориентированы вверх или вниз в зависимости от того, по какую сторону от изоклины они находятся. Форма изоклины хищника, показанная на рис. 7,б. определяется, во-первых, наличием некоторой минимальной плотности жертвы, достаточной для поддержания популяции хищника (при более низкой плотности жертвы хищник не может увеличивать свою численность), а во-вторых, наличием некоторой максимальной плотности самого хищника, при превышении которой численность будет снижаться независимо от обилия жертв.

Рис. 8. Возникновение колебательных режимов в системе хищник-жертва в зависимости от расположения изоклин хищника и жертвы.
При совмещении изоклин жертвы и хищника на одном, графике возможны три различных варианта (рис. 8). Если изоклина хищника пересекает изоклину жертвы в том месте, где она уже снижается (при высокой плотности жертв), векторы, показывающие изменение численности хищника и жертвы, образуют траекторию, закручивающуюся во внутрь, что соответствует затухающим колебаниям численности жертвы и хищника (рис. 8,а). В том случае, когда изоклина хищника пересекает изоклину жертвы в ее восходящей части (т.е. в области низких значений плотности жертв), векторы образуют раскручивающуюся траекторию, а колебания численности хищника и жертвы происходят соответственно с возрастающей амплитудой (рис. 8,б). Если же изоклина хищника пересекает изоклину жертвы в области ее вершины, то векторы образуют замкнутый круг, а колебания численности жертвы и хищника характеризуются стабильной амплитудой и периодом (рис. 8,в).
Иными словами, затухающие колебания соответствуют ситуации, при которой хищник ощутимо воздействует на популяцию жертв, достигнувшую только очень высокой плотности (близкой к предельной), а колебания возрастающей амплитуды возникают, тогда, когда хищник способен быстро увеличивать свою численность даже при невысокой плотности жертв и таким образом быстро ее уничтожить. В других вариантах своей модели Poзенцвейг и Мак-Артур показали, что стабилизировать колебания хищник-жертва можно, введя «убежище», т.е. предположив, что в области низкой плотности жертв существует область, где численность жертвы растет независимо от количества имеющихся хищников.
Стремление сделать модели более реалистичными путем их усложнения проявилось в работах не только теоретиков, но и экспериментаторов. В частности, интересные результаты были получены Хаффейкером (Huffaker), показавшим возможность сосуществования хищника и жертвы в колебательном режиме на примере мелкого растительноядного клеща Eotetranychus sexmaculatus и нападающего на него хищного клеща Typhlodromus occidentalis . В качестве пищи для растительноядного клеща использовали апельсины, помещенные на подносы с лунками (вроде тех, что используются для хранения и перевозки яиц). В первоначальном варианте на одном подносе было 40 лунок, причем в некоторых из них находились апельсины (частично очищенные от кожуры), а в других - резиновые мячики. Оба вида клещей размножаются партеногенетически очень быстро, и поэтому характер их популяционной динамики можно выявить за сравнительно короткий срок. Поместив на поднос 20 самок растительноядного клеща Хаффейкер наблюдал быстрый рост его популяции, которая стабилизировалась на уровне 5-8 тыс. особей (в расчете на один апельсин). Если к растущей популяции жертвы добавляли несколько особей хищника, то популяция последнего быстро увеличивала свою численность и вымирала, когда все жертвы оказывались съеденными.
Увеличив размер подноса до 120 лунок, в которых отдельные апельсины были случайно разбросаны среди множества резиновых мячиков, Хаффейкеру удалось продлить сосуществование хищника и жертвы. Важную роль во взаимодействии хищника и жертвы, как выяснилось, играет соотношение скоростей их расселения. Хаффейкер предположил, что, облегчив передвижение жертвы и затруднив передвижение хищника, можно увеличить время их сосуществования. Для этого на подносе из 120 лунок среди резиновых мячиков располагали случайным образом 6 апельсинов, причем вокруг лунок с апельсинами были устроены преграды из вазелина, препятствовавшие расселению хищника, а для облегчения расселения жертвы на подносе были укреплены деревянные колышки, служившие своего рода «взлетными площадками» для растительноядных клещей (дело в том, что этот вид выпускает тонкие нити и с помощью их может парить в воздухе, распространяясь по ветру). В таком усложненном местообитании хищник и жертва сосуществовали в течение 8 месяцев, продемонстрировав три полных цикла колебаний численности. Наиболее важные условия этого сосуществования следующие: гетерогенность местообитания (в смысле наличия в ней пригодных и непригодных для обитания жертвы участков), а также возможность миграции жертвы и хищника (с сохранением некоторого преимущества жертвы в скорости этого процесса). Иными словами, хищник может полностью истребить то или иное локальное скопление жертв, но часть особей жертвы успеет мигрировать и дать начало другим локальным скоплениям. До новых локальных скоплений хищник рано или поздно тоже доберется, но тем временем жертва успеет расселиться в другие места (в том числе и в те, где она обитала раньше, но потом была истреблена).
Нечто подобное тому, что наблюдал Хаффейкер в эксперименте, встречается и в природных условиях. Так, например, бабочка кактусовая огневка (Cactoblastis cactorum ), завезенная в Австралию, значительно снизила численность кактуса опунции, но не уничтожила его полностью именно потому, что кактус успевает расселиться немного быстрее. В тех местах, где опунция истребляется полностью, перестает встречаться и огневка. Поэтому, когда через некоторое время сюда вновь проникает опунция, то в течение определенного периода она может произрастать без риска быть уничтоженной огневкой. Со временем, однако, огневка снова здесь появляется и, быстро размножаясь, уничтожает опунцию.
Говоря о колебаниях хищник-жертва, нельзя не упомянуть и о циклических изменениях численности зайца и рыси в Канаде, прослеженных по материалам статистики заготовок пушнины компанией Гудзон-Бэй с конца XVIII вплоть до начала XX в. Этот пример нередко рассматривался как классическая иллюстрация колебаний хищник-жертва, хотя на самом деле мы видим только следование роста численности популяции хищника (рыси) за ростом численности жертвы (зайца). Что же касается снижения численности зайцев после каждого подъема, то оно не могло объясняться только возросшим прессом хищников, а было связано с другими факторами, по-видимому, прежде всего нехваткой корма в зимний период. К такому выводу пришел, в частности, М. Джилпин (Gilpin), пытавшийся проверить, могут ли быть описаны эти данные классической моделью Лотки-Вольтерры. Результаты проверки показали, что удовлетворительного соответствия модели нет, но как ни странно, оно становилось лучше, если хищника и жертву меняли местами, т.е. трактовали рысь как «жертву», а зайца - как «хищника». Подобная ситуация нашла свое отражение и в шутливом названии статьи («Едят ли зайцы рысей?»), по сути своей очень серьезной и опубликованной в серьезном научном журнале.
система РА88, которая одновременно предсказывает вероятность более чем 100 фармакологических эффектов и механизмов действия вещества на основе его структурной формулы. Эффективность применения этого подхода к планированию скрининга составляет около 800%, а точность компьютерного прогноза на 300% превосходит предсказание экспертов.
Итак, одним из конструктивных инструментов получения новых знаний и решений в медицине является метод математического моделирования. Процесс математизации медицины – частое проявление взаимопроникновения научных знаний, повышающее эффективность лечебно-профилактической работы.
4. Математическая модель «хищники-жертвы»
Впервые в биологии математическую модель периодического изменения числа антагонистических видов животных предложил итальянский математик В. Вольтерра с сотрудниками. Модель, предложенная Вольтерра, явилась развитием идеи, намеченной в 1924 году А. Лоттки в книге "Элементы физической биологии". Поэтому эта классическая математическая модель известна как модель "Лоттки-Вольтерра".
Хотя в природе отношения антагонистических видов более сложные, чем в модели, тем не менее они являются хорошей учебной моделью, на которой можно изучать основные идеи математического моделирования.
Итак, задача : в некотором экологически замкнутом районе живут два вида животных (например, рыси и зайцы). Зайцы (жертвы) питаются растительной пищей, имеющейся всегда в достаточном количестве (в рамках данной модели не учитывается ограниченность ресурсов растительной пищи). Рыси (хищники) могут питаться только зайцами. Необходимо определить, как будет меняться численность жертв и хищников с течением времени в такой экологической системе. Если популяция жертв увеличивается, вероятность встреч хищников с жертвами возрастает, и, соответственно, после некоторой временной задержки, растет популяция хищников. Эта достаточно простая модель вполне адекватно описывает взаимодействие между реальными популяциями хищников и жертв в природе.
Теперь приступим к составлению дифференциальных уравнений. Обо-
значим число жертв через N, а число хищников через M. Числа N и M являются функциями времени t . В нашей модели учтем следующие факторы:
а) естественное размножение жертв; б) естественная гибель жертв;
в) уничтожение жертв за счет поедания их хищниками; г) естественное вымирание хищников;
д) увеличение числа хищников за счет размножения при наличии пищи.
Так как речь идет о математической модели, то задачей является получение уравнений, в которые входили бы все намеченные факторы и которые описывали бы динамику, то есть изменение числа хищников и жертв со временем.
Пусть за некоторое время t количество жертв и хищников изменится на ∆N и ∆M. Изменение числа жертв ∆N за время ∆t определяется, во-первых, увеличением в результате естественного размножения (которое пропорционально имеющемуся количеству жертв):
где В – коэффициент пропорциональности, характеризующий скорость естественного вымирания жертв.
В основе вывода уравнения, описывающего уменьшение числа жертв изза поедания их хищниками, лежит идея о том, что чем чаще происходит их встреча, тем быстрее уменьшается число жертв. Ясно также, что частота встреч хищников с жертвой пропорционально и числу жертв и числу хищников, то
Поделив левую и правую часть уравнения (4) на ∆t и перейдя к пределу при ∆t→0 , получим дифференциальное уравнение первого порядка:
Для того, чтобы решить это уравнение, нужно знать, как меняется число хищников (М ) со временем. Изменение числа хищников (∆М ) определяется увеличением из-за естественного размножения при наличии достаточного количества пищи (М 1 = Q∙N∙M∙∆t ) и уменьшением из-за естественного вымирания хищников (M 2 = - P∙M∙∆t ):
M = Q∙N∙M∙∆t - P∙M∙∆t |
Из уравнения (6) можно получить дифференциальное уравнение:
Дифференциальные уравнения (5) и (7) представляют собой математическую модель "хищники-жертвы". Достаточно определить значения коэффици-

ентов A, B, C, Q, P и математическую модель можно использовать для решения поставленной задачи.
Проверка и корректировка математической модели. В данной лабора-
торной работе предлагается кроме просчета наиболее полной математической модели (уравнения 5 и 7), исследовать более простые, в которых что-либо не учитывается.
Рассмотрев пять уровней сложности математической модели, можно "почувствовать" этап проверки и корректировки модели.
1-ый уровень – в модели учтено для "жертв" только их естественное размножение, "хищники" отсутствуют;
2-ой уровень – в модели учтено для "жертв" их естественное вымирание, "хищники" отсутствуют;
3-ий уровень – в модели учтены для "жертв" их естественное размножение
и вымирание, "хищники" отсутствуют;
4-ый уровень – в модели учтены для "жертв" их естественное размножение
и вымирание, а также поедание "хищниками", но число "хищников" остается неизменным;
5-ый уровень – в модели учтены все обсуждаемые факторы.
Итак, имеем следующую систему дифференциальных уравнений:
где М – число "хищников"; N – число "жертв";
t – текущее время;
A – скорость размножения "жертв"; C – частота встреч "хищники-жертвы"; B – скорость вымирания "жертв";
Q – размножение "хищников";
P – вымирание "хищников".
1-ый уровень: М = 0, В = 0; 2-ой уровень: М = 0, А = 0; 3-ий уровень: М = 0; 4-ый уровень: Q = 0, Р = 0;
5-ый уровень: полная система уравнений.
Подставляя значения коэффициентов в каждый уровень, будем получать разные решения, например:
Для 3-его уровня значение коэффициента М=0 , тогда

решая уравнение получим
Аналогично для 1-го и 2-го уровней. Что касается 4-го и 5-го уровней, то здесь необходимо решать систему уравнений методом Рунге-Кутта. В результате получим решение математических моделей данных уровней.
II. РАБОТА СТУДЕНТОВ ВО ВРЕМЯ ПРАКТИЧЕСКОГО ЗАНЯТИЯ
Задание 1 . Устно-речевой контроль и коррекция усвоения теоретического материала занятия. Сдача допуска к занятию.
Задание 2 . Выполнение лабораторной работы, обсуждение полученных результатов, оформление конспекта.
Выполнение работы
1. С рабочего стола компьютера вызвать программу "Лаб. №6", щелкнув по соответствующему ярлыку два раза левой клавишей "мыши".
2. Щелкнуть дважды левой клавишей "мыши" по ярлыку "PREDATOR".
3. Выбрать ярлык "PRED" и повторить вызов программы левой клавишей "мыши" (щелкнув дважды).
4. После титульной заставки нажать "ENTER".
5. Моделирование начинать с 1-го уровня.
6. Ввести год, с которого будет проводиться анализ модели: например, 2000
7. Выбрать временные интервалы, например, в течение 40 лет, через 1 год (затем через 4 года).
2-ой уровень: B = 0.05; N0 = 200;
3-ий уровень: A = 0.02; B = 0.05; N = 200;
4-ый уровень: A = 0.01; B = 0.002; C = 0.01; N0 = 200; M = 40; 5-ый уровень: A = 1; B = 0.5; C = 0.02; Q = 0.002; P = 0.3; N0 = 200;
9. Подготовить письменный отчет по работе, который должен содержать уравнения, графики, результаты расчета характеристик модели, выводы по проделанный работе.
Задание 3. Контроль конечного уровня знаний:
а) устно-речевой отчет за выполненную лабораторную работу; б) решение ситуационных задач; в) компьютерное тестирование.
Задание 4. Задание на следующее занятие: раздел и тема занятия, согласование тем реферативных докладов (объем доклада 2-3 стр., регламент 5-7 мин.).
В модели Колмогорова сделано одно существенное допущение: поскольку предполагается, что при то это означает существование в популяции жертв механизмов, регулирующих их численность даже в отсутствие хищников.
К сожалению, подобная формулировка модели не позволяет ответить на вопрос, вокруг которого в последнее время ведется много споров и о котором мы уже упоминали в начале главы: каким образом популяция хищников может оказывать регулирующее воздействие на популяцию жертв с тем, чтобы вся система была устойчива? Поэтому мы вернемся к модели (2.1), в которой механизмы саморегуляции (например, регулирование при помощи внутривидовой конкуренции) в популяции жертв (впрочем, как и в популяции хищников) отсутствуют; поэтому единственным механизмом регуляции численностей, входящих в сообщество видов, являются трофические взаимоотношения между хищниками и жертвами.
Здесь (так что, в отличие от предыдущей модели, Естественно, что решения (2.1) зависят от конкретного вида трофической функции которая, в свою очередь, определяется характером хищничества, т. е. трофической стратегией хищника и защитной стратегией жертвы. Общими для всех этих функций (см. рис. И) являются следующие свойства:
Система (2.1) имеет одну нетривиальную стационарную точку, координаты которой определяются из уравнений
![]()
при естественном ограничении .
Существует еще одна стационарная точка (0, 0), соответствующая тривиальному равновесию. Нетрудно показать, что эта точка - седло, а сепаратрисами являются координатные оси.
Характеристическое уравнение для точки имеет вид
![]()
Очевидно, что для классической вольтерровской модели .
Поэтому величину f можно рассматривать как меру отклонения рассматриваемой модели от вольтерровской.
![]()
стационарная точка - фокус, и в системе появляются колебания; при выполнении противоположного неравенства - узел, и колебания в системе отсутствуют. Устойчивость этого состояния равновесия определяется условием
т. е. существенным образом зависит от вида трофической функции хищника.
Условие (5.5) можно интерпретировать следующим образом: для устойчивости нетривиального равновесия системы хищник - жертва (и тем самым, для существования этой системы) достаточно, чтобы в окрестности этого состояния относительная доля потребляемых хищником жертв возрастала с возрастанием численности жертв. Действительно, доля жертв (от общей их численности), потребляемых хищником, описывается дифференцируемой функцией условие возрастания которой (положительность производной) выглядит как
![]()
Последнее условие, взятое в точке есть не что иное, как условие (5.5) устойчивости равновесия. При непрерывности оно должно выполняться и в некоторой окрестности точки Таким образом, если численности жертв в этой окрестности, то
Пусть теперь трофическая функция V имеет вид, изображенный на рис. 11, а (характерный для беспозвоночных). Можно показать, что для всех конечных значений (поскольку выпукла вверх)
т. е. при любых величинах стационарной численности жертв неравенство (5.5) не выполняется.
Это означает, что в системе с таким видом трофической функции не существует устойчивого нетривиального равновесия. Возможны несколько исходов: либо численности как жертвы, так и хищника неограниченно возрастают, либо (при прохождении траектории вблизи одной из координатных осей) в силу случайных причин численность жертвы или численность хищника станет равной нулю. При гибели жертвы через некоторое время погибнет и хищник, если же сначала погибает хищник, то численность жертвы начнет экспоненциально возрастать. Третий вариант - возникновение устойчивого предельного цикла - невозможен, что легко доказывается.
В самом деле, выражение

в положительном квадранте всегда положительно, если только имеет вид, изображенный на рис. 11, а. Тогда, согласно критерию Дюлака в этой области отсутствуют замкнутые траектории и устойчивого предельного цикла существовать не может.
Итак, можно сделать вывод: если трофическая функция имеет вид, изображенный на рис. 11, а, то хищник не может быть регулятором, обеспечивающим стабильность популяции жертвы и тем самым устойчивость всей системы в целом. Система может быть устойчива только в том случае, когда в популяции жертвы существуют собственные внутренние регулирующие механизмы, например, внутривидовая конкуренция или эпизоотии. Этот вариант регулирования был уже рассмотрен в §§ 3, 4.
Ранее отмечалось, что такой тип трофической функции характерен для хищников-насекомых, «жертвами» которых также обычно являются насекомые. С другой стороны, наблюдения за динамикой многих природных сообществ типа «хищник - жертва», включающих в себя виды насекомых, показывают, что для них характерны колебания очень большой амплитуды и весьма специфического вида.
Обычно после более или менее постепенного роста численности (который может происходить либо монотонно, либо в виде колебаний с возрастающей амплитудой) происходит ее резкое падение (рис. 14), и затем картина повторяется. По-видимому, такой характер динамики численностей видов насекомых может быть объяснен неустойчивостью этой системы при малых и средних значениях численности и действием мощных внутрипопуляционных регуляторов численности при больших ее значениях.

Рис. 14. Динамика численности популяции австралийской листоблошки Cardiaspina albitextura, кормящейся на эвкалиптах. (Из статьи: Clark L. R. The population dynamics of Cardiaspina albitextura.-Austr. J. Zool., 1964, 12, № 3, p. 362-380.)
Если же в систему «хищник - жертва» входят виды, способные к достаточно сложному поведению (например, хищники способны к обучению или жертвы способны находить убежище), то в такой системе возможно существование устойчивого нетривиального равновесия. Доказывается это утверждение весьма просто.
В самом деле, трофическая функция тогда должна иметь вид, изображенной на рис. 11, в. Точка на этом графике - точка касания прямой, проведенной из начала координат, графика трофической функции Очевидно, что в этой точке функция имеет максимум. Также легко показывается, что для всех выполняется условие (5.5). Следовательно, нетривиальное равновесие, в котором численность жертв меньше будет асимптотически устойчиво
Однако мы ничего не можем сказать о том, сколь велика область устойчивости этого равновесия. Например, если существует неустойчивый предельный цикл, то эта область должна лежать внутри цикла. Или другой вариант: нетривиальное равновесие (5.2) неустойчиво, но существует устойчивый предельный цикл; в этом случае также можно говорить о стабильности системы хищник - жертва. Поскольку выражение (5.7) при выборе трофической функции типа рис. 11, в может менять знак при изменении при , то критерий Дюлака здесь не работает и вопрос о существовании предельных циклов остается открытым.
